Carder
Professional
- Messages
- 2,619
- Reaction score
- 1,877
- Points
- 113

Гесперорнис, птица позднего мелового периода, имела крылья, похожие на крылья археоптерикса.
Гарри Говьер Сили разделил динозавров на две группы: отряд Saurischia (динозавры с «ящерицами») и отряд Ornithischia (динозавры с «птичьими шипами»). У обоих отрядов, вероятно, был общий предок, живший где-то в среднем триасе. Птицы принадлежат к кладе заурисховых динозавров.
Как и у всех наземных животных, на каждой стороне таза было по три кости. Левая и правая подвздошные кости (единственное число: ilium) прочно сжимали позвоночник в крестце. Левая и правая лобковые кости (единственное число: лобок) простираются ниже подвздошной кости. Левая и правая седалищные кости (единственное число: седалищная кость) простираются вниз и назад под подвздошной костью и позади лобка. У некоторых динозавров лобковые волосы вытянуты вниз и вперед, как у ящериц. Вот почему Сили назвал их заурисхийскими, или «ящеричными» динозаврами. У других динозавров лобковые волосы вытянуты вниз и назад, проходят под седалищной костью и параллельно ей, как у птиц. Сили назвал этих динозавров орнитисхийскими, или «птичьими» динозаврами.
Таз орнитисхиевых динозавров (множественное число от таза) развился в результате эволюционных изменений примитивного заурисхиевого таза. У орнитизхов были и другие черты, объединяющие их: кость, называемая предзубной, находилась в передней части нижней челюсти. Кроме того, верхняя часть глазницы обрамляла кость «веко».
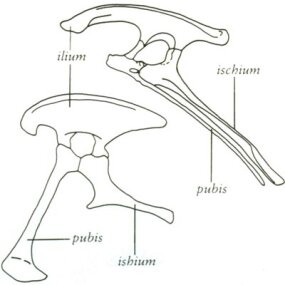
Верхний таз типичен для орнитисхийных, или «птичьих» динозавров. Дно типично для заурисхийных, или «ящеричных» динозавров.
Некоторые динозавры не были ни заурисхами, ни орнитисхами. Самые ранние и примитивные динозавры, такие как Стаурикозавр и Герреразавр, не вписываются ни в один из этих порядков. Они были слишком специализированы, чтобы быть прямыми предками динозавров.
СОДЕРЖАНИЕ
- Заурисхии: Sauropodomorpha
- Заурисхия: Theropoda
- Орнитисция: Тинеофора
- Орнитисция: Marginocephalis
- Орнитисция: Ornithopoda
Заурисхии: Sauropodomorpha

Sauropelta, бронированный динозавр раннего мелового периода.
Зауроподоморфы быстро эволюционировали в две основные группы: Prosauropoda и Sauropoda. Хотя прозауроподы появились раньше, ни один известный прозауропод не мог быть предком зауроподов. Прозавроподы были широко распространены и насчитывали не менее семи семейств. Они жили до ранней юры. Самые большие прозауроподы, некоторые из которых достигли 40 и более футов, были динозаврами с прямыми конечностями, в чем-то напоминающими более поздних зауроподов. Все прозауроподы питались растениями.
Семейство: Thecodontosauridae: Самый примитивный прозауропод, Thecodontosaurus, был также одним из самых маленьких. Его длина была от шести до десяти футов. Как у всех прозауроподов и большинства зауроподов, у него были выдающиеся когти на каждой передней лапе и большие когти на каждой задней.
Семейство: Plateosauridae: это самое известное семейство прозауроподов, животные обитают в Европе, Китае, Северной и Южной Америке. Они были от 25 до 30 футов в длину, с узкой длинной мордой, длинной шеей, мощными передними и задними конечностями и тяжелым телом.
Инфраотряд: Sauropoda
Вторая группа зауроподоморфов, зауроподы, вероятно, произошла от предка, очень похожего на Thecodontosaurus. Вероятно, это произошло где-то в позднем триасе, когда впервые появились зауроподы. Все зауроподы были гигантами и четвероногими растительноядными.
Как и современные слоны, зауроподы мало боялись хищников из-за своего размера. Большой размер также помогал им дотянуться до еды, такой как листья на верхушках деревьев, которая была слишком высокой для более мелких поедателей растений. У зауроподов было много особенностей из-за их большого размера. Они потеряли хватательную функцию передних ног, и их ноги стали похожи на длинные прямые колонны. Их позвонки (кости в позвоночнике) имели глубокие впадины, чтобы облегчить вес их позвоночника. Кроме того, для увеличения силы у них было больше позвонков в местах соединения таза и позвоночника.
Черепа зауроподов были либо тупыми (плоскими), либо сужающимися (заостренными), а ноздри были отклонены от кончика морды. Головы зауроподов, которые были легко сложены и хрупки, часто отламывались после смерти.
Семейство: Vulcanodontidae: Самый ранний настоящий зауропод - это Vulcanodon из ранней юры Зимбабве. Единственный скелет лишен головы, шеи и большей части хвоста. У него было массивное тело и длинные прямые ноги. Передние конечности были почти такой же длины, как и задние, а на каждой задней ноге было по пять пальцев.
Семейство: Barapasauridae: Следующий по первобытности зауропод, Barapasaurus, известен по частям нескольких скелетов ранней юры Индии. Он был до 60 футов в длину, с тонким телом, длинной шеей, хвостом и конечностями.

Omeisaurus tianfuensis, семейство Euhelopodidae.
Семейство: Euhelopodidae: большинство зауроподов, известных из средней и поздней юры Китая, теперь помещено в отдельное семейство, Euhelopodidae. Euhelopodids - одно из наиболее примитивных семейств зауроподов, но они включают таких экзотических животных, как Mamenchisaurus с очень длинной шеей и Omeisaurus.
Семья: Cetiosauridae: это семейство из средней юры, возможно, от предка из семейства Vulcanodontidae. Цетиозавриды распространились и распространились в Европе, Северной и Южной Америке, Африке и Австралии к средней юре.
Черепа у цетиозавров были тупыми и прямоугольными, с ноздрями сбоку от морды. Шея была короткой, обычно с 12 позвонками. У зауроподов они варьировались от маленьких до больших; большинство из них были от 35 до 60 футов в длину. Самый известный род - шунозавр из средней юры Китая. На конце хвоста у него была небольшая костлявая дубинка. Цетиозавриды просуществовали до поздней юры.
Семейство: Brachiosauridae: передние конечности брахиозаврид были такими же или длиннее задних конечностей. Это придавало телу наклон назад от шеи к хвосту. Число шейных позвонков в этом семействе увеличилось до 13 и более. Ноздри находились дальше от кончика тупой морды и над глазами в черепе брахиозавра. Большинство брахиозаврид были крупнее цетиозавридов, 80 или более футов в длину, хотя у них были более короткие хвосты. Они были одними из самых тяжелых известных наземных животных. Чтобы уменьшить вес, их огромные позвонки были почти полностью полыми. Известные во всем мире брахиозавриды появляются в летописи окаменелостей в течение средней юры, были наиболее многочисленны в поздней юре и почти исчезли к концу раннего мелового периода.
Семейство: Camarasauridae: В этом семействе череп был коробчатым. У них осталось 12 шейных позвонков, а передние конечности были немного короче задних. Одним из последних известных камаразаврид был Opisthocoelicaudia из Монголии, массивный зауропод с коротким хвостом, который, вероятно, помогал ему, когда он стоял на задних конечностях, чтобы достать пищу.
Семейство: Diplodocidae: это семейство включает некоторых из самых известных зауроподов, в том числе апатозавра и диплодока. Черепа диплодоцидов были длинными, сужающимися к ложкообразной морде и имели ноздри на вершине черепа. Их маленькие стержневидные зубы находились в передней части морды. Диплодоциды имели длинные шеи, до 15 позвонков. Их спины были короткими по сравнению с длиной их задних конечностей, а хвосты заканчивались хлыстовой травмой, которая, вероятно, использовалась в качестве оружия. Длинные шеи и хвосты сделали некоторых диплодоцидов самыми длинными животными из когда-либо существовавших.
Семейство: Titanosauridae: Почти все зауроподы южного полушария из позднего мела и многие более ранние были титанозавридами. Их конечности были коренастыми. Позвонки передней и средней части хвоста уникальны и являются лучшей чертой, которая отличает семейство. Не было найдено ни одного полного или почти полного черепа титанозаврида. Одним из самых интересных титанозавров был сальтазавр, который был приземистым и покрыт броней, похожей на анкилозавры.
Большинство титанозавридов было от 40 до 50 футов в длину, но некоторые из них стали гигантскими. Титанозавриды жили в основном в южном полушарии в меловой период, выживая там, когда зауроподы в северном полушарии вымерли.
Заурисхия: Theropoda

Целофиз.
Все тероподы были хищными динозаврами, кроме герреразавров. От самых маленьких динозавров до крупнейших мясоедов, у теропод были самые разные виды заурисхических динозавров из всех подотрядов. У этих двуногих мясоедов были когтистые лапы с не более чем тремя функциональными пальцами.
Крылья и лапы птиц похожи на руки и ноги скелетов теропод. Как и птицы, у всех теропод в той или иной степени были полые кости. Лучшая птица-предок - это маленький пернатый тероподоподобный археоптерикс из поздней юры.
Тероподы разделились на две основные группы: Ceratosauria с гибкими хвостами; и столбняк с жесткими хвостами. Все самые ранние тероподы были цератозаврами. Их летопись окаменелостей относится к периоду от позднего триаса до поздней юры. Тетануранцы появились в средней юре, разнообразились в поздней юре и были основными хищниками северного полушария до позднего мела.
Инфраотряд: Ceratosauria
Семейство: Podokesauridae: Самые ранние цератозавры включают целофизов из позднего триаса на западе Северной Америки. Он был маленьким и шустрым, с длинным тонким черепом с множеством зубов.
Семейства: Halticosauridae и Ceratosauridae: Дилофозавр, который жил в ранней юре, имел двойной гребень на голове. Цератозавр был из позднего юрского периода и имел на голове рог. Оба были из Северной Америки и являются примерами более поздних представителей цератозавров. После поздней юры цератозавры, по-видимому, исчезли в северном полушарии, но выжили в Южной Америке.
Семейство: Abelisauridae: Абелизавриды - это группа средних и крупных африканских и южноамериканских теропод, для которых характерны короткие и высокие черепа. Карнотавр из Аргентины и Маджунгатолус из Мадагаскара похожи, за исключением того, что у Карнотавра есть два больших рога на черепе.

Abelisaurus comahuensis.
Инфраотряд: тетануры
Столбняки, наиболее продвинутые теропод, включали несколько групп, взаимоотношения которых не совсем понятны. На голове обычно не было гербов и других украшений. Их руки имели три или меньше пальцев, а у «большого пальца» обычно был самый большой коготь.
Семейство: Compsognathidae: Самым примитивным тетанураном был Compsognathus из поздней юры Европы. Это был самый маленький теропод, около трех футов длиной и легкого сложения.
Семейство: Coeluridae: Ornitholestes и Coelurus, которые жили в поздней юре в западной части Северной Америки, были быстрыми, легко сложенными тероподами, достигающими двух-трех футов высотой в бедрах и длиной от шести до десяти футов.
Семейство: Carcharodontosauridae: Эта группа гигантских теропод из Гондваны включает огромных хищных динозавров, гиганотозавров из Аргентины и Carcharodontosaurus из Северной Африки.
Семейство: Therizinosauridae: Теризинозавриды, по- видимому, были травоядными или всеядными тероподами, известными из позднего мела в Азии и Северной Америке. Необычный птицеподобный таз и почти прозауроподный череп теризинозаврид привели к неопределенности в отношении их эволюционного положения, но недавно было показано, что они являются тероподами, тесно связанными с орнитомимидами.
Семейство: Spinosauridae: Спинозавриды - отличительная группа теропод с длинными крокодилоподобными мордами и удлиненными позвоночными шипами, которые могли образовывать на спине паруса. Спинозавриды встречаются только в меловом периоде, но известны из Африки, Южной Америки и Европы.
Семейство: Oviraptorisauridae: Еще одним любопытным тероподом из мелового периода Азии является Oviraptor, у которого высокий, очень пневматический череп с клювом, похожим на черепаху. Овираптор получил свое название («яичный хищник») потому, что в Монголии были обнаружены экземпляры, которые изначально считались гнездами яиц цератопсов.
Семейство: Allosauridae: это семейство типично для более крупных теропод юрского и раннемелового периода, длина которых составляла от 15 до 35 футов или больше. Самый большой аллозаврид мог быть более 40 футов в длину. Аллозавриды были стройными, но опасными хищниками.
Семейство: Tyrannosauridae: Самыми драматичными из всех теропод были тираннозавриды, которые, вероятно, произошли от аллозавридоподобных предков в начале позднего мелового периода. В отличие от других тетануранов, у них были массивные тела, головы необычной формы и маленькие двупалые руки.

Тираннозавр нападает на Эдмонтозавра.
Некоторые тираннозавриды были почти 50 футов в длину и стали самыми крупными из известных мясоедов на суше. Самые маленькие, такие как Nanotyrannus, были около 18 футов в длину. Тираннозавриды среднего размера, такие как альбертозавр и дасплетозавр, были около 30 футов в длину.
Семейство: Ornithomimidae: В меловом периоде также возникли многие виды мелких теропод. Их скелеты были похожи на птиц. У большинства были изменены передние ноги и руки для мощного удара. «Быстрый удар» движения передних конечностей мог быть началом мощного взмаха крыльев птиц.
«Страусиные динозавры» или Ornithomimidae известны лучше всего из позднего мела Восточной Азии и западной части Северной Америки. У них были маленькие головы и обычно не было зубов. У них были длинные шеи и короткие жесткие спины. Их передние конечности были длинными, а мощные задние были созданы для скорости. Считается, что они были самыми быстрыми динозаврами.
Семейство: Dromaeosauridae: Дейноних и другие теропод с «серповидным когтем» - одни из наиболее изученных динозавров. Открытие Дейнониха поддержало идею взаимоотношений между птицами и динозаврами и положило начало дебатам о теплокровных динозаврах. У каждой ступни на втором пальце был большой серповидный коготь. На конце хвоста были позвонки, которые сцеплялись вместе, чтобы сделать его жестким.
Семейство: Troodontidae: Эта группа теропод с «серповидными когтями» имела большой мозг и большие глаза, обращенные вперед. Некоторые, такие как Troodon и Saurornithoides, могли быть почти такими же умными, как некоторые млекопитающие.
Орнитисция: тинеофора
Дацентрурус Арматус.
Самым ранним динозавром-орнитисханом был Пизанозавр, двуногий (двуногий) растительноядный двуногий трехфутовый в длину, живший в конце среднего триаса в Аргентине. Все орнитишианы питались растениями.
Позднее орнитисхианы разделились на три продвинутые группы: тяжелые, бронированные едоки растений, передвигающиеся на четвереньках; специализированные куполообразные динозавры и рогатые динозавры; и двуногие едоки растений, в том числе игуанодон и утконосые динозавры.
Семейство: Fabrosauridae: это семейство встречается в породах от позднего триаса до поздней юры на нескольких континентах. К этому семейству обычно относятся и другие примитивные орнитисхианы. Он был назван в честь Фаброзавра из ранней юры Южной Африки. Все фаброзавриды были небольшими двуногими растительноядными. Самый известный фаброзаврид - Лесотозавр, тоже из ранней юры на юге Африки. Таз лесотозавра имеет некоторые особенности, которые ставят его в основу орнитизхий. Скутеллозавр из ранней юры западной части Северной Америки был защищен небольшими костными пластинами, напоминающими большие пластины более поздних бронированных орнитисхий.
ТИРЕОФОРА.
Инфраотряд: Стегозаврия
Стегозавры были главными бронированными динозаврами юрского периода; анкилозавры остались в тени. Но анкилозавры заменили стегозавров в меловой период. Скелидозавр был очень похож на обе группы. Он был из ранней юры Англии и ходил на всех четырех конечностях (четвероногий). У него были зубы, подобные стегозавру, узор из пластин и шипов, напоминающий анкилозавра, и таз, как у скутеллозавра.
Семейство: Stegosauridae: эти бронированные динозавры, вероятно, эволюционировали в Китае во время ранней юры. К позднему юрскому периоду они были в Европе (Дацентрурус), Северной Америке (Стегозавр) и Африке (Кентрозавр).
Доспех представлял собой двойной ряд больших костных пластин, тянувшихся вдоль спины от задней части головы до хвоста. Острые шипы на конце хвоста использовались как оружие против хищников.
Инфраотряд: анкилозаврия
В раннем меловом периоде анкилозавры вытеснили стегозавров повсюду, кроме Индии. Анкилозавры отличались от стегозавров тем, что у них был гибкий нательный доспех, а не двойной ряд высоких пластин. К тому же они были ближе к земле, с небольшим изгибом спины, если таковой был. Головы стегозавров были длинными и узкими, но у анкилозавров были короткие, дощатые черепа. Вместо хвостовых шипов у анкилозавров для защиты были плечевые шипы или хвостовые булавы.
Семейство: Nodosauridae: наиболее примитивные анкилозавры, включая все роды юрского периода и южного полушария, принадлежат к этому семейству. У некоторых нодозавридов были большие конические шипы на шее и плечах для защиты.
Семейство: Ankylosauridae: это семейство могло возникнуть в раннем меловом периоде от предка нодозаврид. У черепов анкилозаврид были рога, выступающие сзади, что придавало им треугольную форму, если смотреть сверху. У всех анкилозавридов для защиты были массивные булавы с костлявым хвостом.
Орнитисция: Marginocephalis
Homocephale calathocercos.
Яверландия, родом из раннего мелового периода Англии, является старейшим из известных пахицефалозавров. Но единственный известный образец - это маленькая толстая тюбетейка с двумя костяными бугорками, которые мало что говорят о том, как могло выглядеть животное или его взаимоотношения. Куполообразные динозавры были немногочисленными животными мелового периода от мелких до средних. Большинство из них обитало в северном полушарии, но один род, Majungatholus, известен из позднего мела на Мадагаскаре.
Задняя часть головы куполообразных динозавров была расширена до выступа, на котором часто были костлявые бугорки или короткие шипы. У одного семейства, Homalocephalidae, кости на вершине черепа были плоскими и толстыми (как у Homalocephale и Goyocephale из Монголии).
В другом семействе, Pachycephalosauridae, кости были подняты в очень толстый высокий купол, который был главной особенностью внешнего вида животного, и даже вырастали над выступом (как у Stegoceras). Скелеты пахицефалозавров редки, но их черепа-купола, поскольку они были твердой костью, часто были окаменелыми. Кое-где они встречаются довольно часто.
У пахицефалозавров были широкие пухлые тела. Они были двуногими травоядными. По мере того, как животное росло, его купол становился все больше. Толстые купола использовались, чтобы соревноваться за самцов в схватках между самцами бодами головой или для борьбы с хищниками, бодая их боком.
Инфраотряд: Цератопсия
Цератопсии были рогатыми динозаврами и их родственниками. Они отличались от пахицефалозавров - и всех других динозавров - тем, что у них была особая кость, ростральная, которая составляла часть большого, похожего на попугая клюва.
Семейство: Psittacosauridae: самые старые и примитивные цератопсы принадлежат к семейству небольших двуногих бегунов, таких как Psittacosaurus из раннего мелового периода Китая и Монголии.
Семейство: Protoceratopsidae: в этом семействе задняя часть черепа была расширена до широкой оборки на задней части шеи. Микроцератопс был двуногим животным, как и его возможные предки пситтакозавриды. Другие протоцератопсиды ходили на четвереньках, что облегчало поддержку их больших голов.
Семейство: Ceratopsidae: у Ceratopsidae был самый короткий ареал среди всех семейств динозавров. Они возникли в позднем меловом периоде на западе Северной Америки. Они быстро превратились во множество необычных форм и жили до конца мезозойской эры. Четвероногие (четвероногие) цератопсиды размером от коровы до слона имели рога и оборки на голове.

Arrhinoceratops brachyops.
У них были мощные челюсти с сотнями зубов для срезания жестких растений. У трицератопса были самые мощные челюстные мышцы среди всех наземных животных. Рога использовались как оружие. Оборки могли защищать шею и были ярко окрашены для брачного сезона. Оборки, возможно, также помогли сохранить температуру его тела.
Орнитисция: Ornithopoda

Гетердонтозавр tucki.
За исключением увеличения размеров и эволюции самого замечательного жевательного устройства среди всех динозавров, разные семейства орнитоподов отличались друг от друга в незначительных деталях. Все были двуногими травоядными.
Семейство: Heterodontosauridae: Эти маленькие, подвижные двуногие едоки растений были обнаружены в основном в скалах ранней юры на юге Африки.
Их зубы были острыми и напоминали клыки спереди, но зубы по бокам челюсти были созданы для жевания и резки растений. У них были большие клыкоподобные зубы (конусообразные, заостренные зубы) по углам верхней и нижней челюстей.
Семейство: Hypsilophodontidae: это было наиболее распространенное и долгоживущее семейство орнитопод. Он процветал почти во всем мире от средней юры до конца мелового периода.
Гипсилофодонтиды были небольшими, но имели относительно большие головы. Их ступни были примитивными, с четырьмя функциональными пальцами. Первый палец ноги был меньше остальных; пятый палец был всего лишь шиной. У гипсилофодонтидов были маленькие передние конечности с крошечными руками. Костные сухожилия укрепляли спину и жесткость хвоста. Большинство из них были маленькими динозаврами длиной от шести до десяти футов, но некоторые виды тенонтозавров достигали 22 футов в длину. Поздний меловой тескелозавр вырос примерно до 18 футов в длину.
Семья: Dryosauridae: это недолговечное семейство возникло примерно в то же время, что и Hypsilophodontidae. Самым ранним дриозавридом был дриозавр из поздней юры западной части Северной Америки и восточной Африки. Валдозавр из раннего мела Европы и Северной Африки и кангназавр из Африки - два других рода в этом семействе. Дриозавриды с маленькими передними конечностями и головами были крупнее и мощнее гипсилофодонтидов. У них не было зубов на передней части морды, а вместо этого был хорошо развит клюв, который мог иметь роговой покров.
Семейство: Camptosauridae: позднеюрский род Camptosaurus из западной части Северной Америки был пухлым орнитоподом среднего размера около 15 футов в длину. У него были особые ступни и череп.
Семейство: Iguanodontidae: Игуанодон - один из самых известных динозавров. У этого громоздкого орнитопода 35 футов длиной был глубокий узкий череп; крепкий, хорошо развитый таз; вдоль спины проходят ряды костных сухожилий; рука, в которой большой палец превратился в острый шип; и три широких пальца плюс внутренний палец, который превратился в шину. Зубы были толстыми и постоянно заменялись. Он ел жесткие растения, игуанодон ходил на двух ногах, но он также мог ходить на четвереньках руками.

Ранний рисунок (1895 г.) игуанодона.
Игуанодонтиды были обнаружены в формациях раннего и позднего мела в Северной Америке, Северной Африке, Европе и Азии. Наиболее продвинутыми были пробактрозавр из Центральной Азии и уранозавр из Северной Африки. Зубы пробактрозавра заменялись более сложным образом, чем зубы игуанодона. Позвонки уранозавра имели очень длинные шипы, образующие парус. С появлением утконосых динозавров игуанодонтиды исчезли.
Семейство: Hadrosauridae: Эта группа состоит из двух групп, Hadrosaurinae и Lambeosaurinae. Оба они были близкими родственниками утконосых динозавров. Они были крупными и очень крупными пожирателями растений в позднем меловом периоде Северной и Южной Америки и Евразии. Самыми крупными были шантунгозавр из Китая и Lambeosaurus laticaudus из Мексики, оба из которых, возможно, вырастали более чем на 50 футов в длину и весили более 20 тонн. Возможно, они были самыми крупными из известных животных, которые могли ходить на двух ногах. Большинство утконосых динозавров были около 30 футов в длину.
Зубы располагались в 30-40 вертикальных рядах, как ступеньки движущегося эскалатора. По мере того, как каждый зуб стирался, его заменял зуб прямо под ним. Этот процесс начался, когда (или даже раньше) животное вылупилось, и продолжалось до тех пор, пока оно было живым. Черепа гадрозавринов обычно были длиннее и не такими глубокими, как у ламбеозавринов, а их утиные клювы были более плоскими и широкими.
У некоторых гадрозавринов отсутствовали черепные гребни (Edmontosaurus); у других были дугообразные носовые кости (Kritosaurus); у других были твердые черепные гребни (Prosaurolophus и Maiasaura). Но у всех ламбеозавров, черепа которых хорошо известны (гипакрозавр, коритозавр, ламбеозавр и паразауролоф), были полые гребни с петляющими носовыми ходами, которые, возможно, использовались для создания звуков.
Утиные динозавры были, пожалуй, самыми продвинутыми из всех динозавров. У них был отличный слух, зрение, голос и обоняние. Они жили огромными стадами и, возможно, мигрировали сезонно, возвращаясь в одно и то же место каждый год, чтобы спариваться и откладывать яйца.